Diadema antillarum, commonly known as the long-spined black urchin, is classified as follows (Harvey 1956):
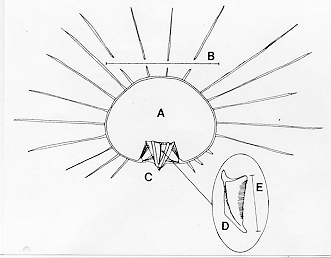
An urchin's body size is measured by test diameter and spine length. The test is the shell that contains the central part of the body (excluding the spines), shaped like a flattened sphere. A diagram of these body parts is given in Figure 1. An average adult individual of this species, as pictured in Figure 2, has a test diameter of about 5 cm and spines measuring at least three times that length (Kaplan 1982). Some individuals have spines measuring up to 30 cm, and the 3:1 ratio of spine length to test diameter still holds.
Diadema antillarum's feeding habits have made this species both partner and predator of corals. Its feeding apparatus, as seen in Figure 1, consists of five strong teeth guided through individual pyramids on the underside of the urchin, and these five pyramids together make up Aristotle's lantern (Levitan 1992). This extendable chewing assembly gives D. antillarum great scraping power, which is very useful in grazing algae and an occasional invertebrate from the coral surface (Bak & van Eys 1975). The scraping action of these teeth not only removes digestible food for the urchin, but also removes some of the coral substrate, thus making D. antillarum an important bioeroder on the reefs (Lessios 1988a).
Diadema antillarum's foods are varied, but it prefers turf algae, which grow abundantly on corals when left unchecked (Carpenter 1981). When turf algae are unavailable, D. antillarum will settle for other types of algae, or may scavenge the sandy sea floor for detritus. This species has even been known to eat live coral organisms when the favored algae are in short supply (Bak & van Eys 1975). Therefore, D. antillarum, formerly described as exclusively herbivorous, is more accurately an omnivorous grazer, and a facultative predator on living corals. However, because it does prefer algae (Carpenter 1981), and because its herbivory is considered a primary factor in its competitive interactions with other reef animals, I will at some points refer to D. antillarum as a herbivore.
In varying urchin population densities, Diadema antillarum can play an important role in coral abundance and community structure. At very low Diadema densities, algae tend to monopolize substrate surface area, effectively preventing coral settlement and causing mortality of adult coral organisms (Sammarco 1980). On the other hand, excessively high densities of D. antillarum can also cause coral mortality and stifle recruitment by abrasive grazing on coral substrate (Sammarco 1980). As mentioned before, significant grazing on coral organisms occurs primarily in food-limited populations of high density (Bak & van Eys 1975), so the ideal conditions for recruitment and growth of live corals include intermediate densities of D. antillarum (Sammarco 1980).
Diadema antillarum was, until 1983, a ubiquitous and ecologically significant herbivore on coral reefs of the Caribbean. Pre-mortality population densities as high as 71 per m2 have been recorded (Sammarco 1982a). D. antillarum has been described by many (e.g., Sammarco 1982a & 1982b; Carpenter 1981) as a main herbivore on Caribbean reefs, and is even considered by some (Lessios 1988a) to have been the most important bioeroder of the reef framework.
The favorite habitat of D. antillarum is the shallow coral reef, especially in depths of 15 m or less. Beyond 10 m deep, urchin population density and grazing intensity decrease with depth (Morrison 1988). Studies in different regions of the same reefs in different seasons have indicated that this urchin prefers to feed in lighter water movement (Foster 1987), and thus may retreat to the more peaceful backreef area (lagoon) during periods of vigorous wave activity.
A major part of D. antillarum's role in coral reef ecosystems involves its interactions with reef fishes. It competes for food with several herbivorous fish species, especially members of the families Acanthuridae (surgeonfishes) and Scaridae (parrotfishes). According to Lewis & Wainwright (1985), acanthurids predominate in shallow areas (<5 m), while scarids are dominant in deeper areas. Diadema exclusion experiments have suggested that this urchin is an excellent competitor for food, as indicated by tremendous changes in algal species composition and in herbivorous fish population density and grazing intensity (Foster 1987; Morrison 1988; Sammarco et al. 1974). At the time when this thesis was written, no data were available on Diadema response to herbivorous fish exclusion.
Another important type of fish-urchin interaction is predation. According to Hunte et al. (1986), at least 15 species of fish prey on D. antillarum, most commonly members of the families Batrachoididae (toadfishes), Balistidae (triggerfishes), Labridae (wrasses), and Pomadasyidae (grunts). This relationship of predators to sea urchins to algae has been compared to the otter-urchin-kelp relationship that exists in temperate kelp forests of the Pacific (Carpenter 1984; Duggins 1980).
Diadema antillarum's reproductive strategy has also been the subject of much scrutiny since the mass mortality. Gametogenesis, fertilization, recruitment, and growth are all essential parts of restoring, or even maintaining, a population. In all these categories, D. antillarum seems to have the preferable strategy for recovery.
Gametogenesis in each individual is not influenced by the size of the population in which it lives (Lessios 1988b). Fertilization takes place externally, as sperm and eggs are released into the water during lunar-synchronized spawnings. The planktonic eggs are fertilized in the water to produce free-swimming larvae, which are then recruited at various reefs, often many miles from their parents' home (Lessios 1988b). While it may seem at first that external fertilization of planktonic gametes would be difficult in reduced population densities, it is important to note the lunar synchrony of aggregation and spawning, which brings even the few existing adults into sufficient proximity to achieve fertilization.
However, it seems that the quantities of planktonic larvae being produced are insufficient to sustain a significant level of recruitment, at least in Panamá (Lessios 1988b). The fact that juveniles were recruited to 1 - 1.5 cm immediately after the mass mortality supports the hypothesis that recruitment is not dependent on the presence and protection of adults (Lessios 1988b). In contrast, juveniles of the Pacific urchin Strongylocentrotus franciscanus do depend on adults for protection during the early stages of growth (Tegner & Dayton 1975). Unfortunately, not much information is available as to what factors limit the viability of planktonic D. antillarum larvae, so it is not yet clear where the large numbers of larvae go. The relative importances of predation, food limitation, and inhospitable environment at this crucial life stage have not yet been adequately determined.
The density independence of recruitment in D. antillarum is very important to the recovery of a species in which most of the large adults have been wiped out. Furthermore, because growth in individuals of this species seems to be inversely related to population density, urchins existing in post-mortality densities may grow faster and reach sexual maturity sooner (Hunte et al. 1986). While it may take D. antillarum several decades (or longer) to return to pre-mortality population density, the increase in body size may re-establish urchin biomass and suppress algal biomass to pre-mortality levels much sooner (Levitan 1988a). The wide dispersal of larvae, along with the independence of recruitment and mortality from local population density, suggests that local populations of D. antillarum are open and recruitment-limited (Karlson & Levitan 1990). This will have important implications for any attempt at restoring these populations to anything resembling their former magnitude.
{kind=link}
{kind=link}